|
Нейрон Собственной Персоной
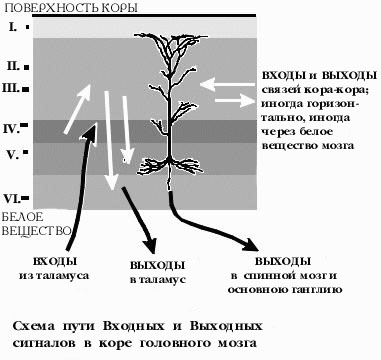
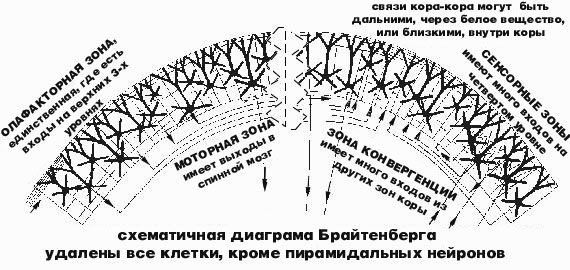
"СЕРОЕ" ВЕЩЕСТВО КОРЫ ГОЛОВНОГО МОЗГА представляет собой всего лишь тонкую корочку на поверхности белого вещества. Но внутри этой красно-коричневой глазури для белого пирога можно различить нейроны, организованные в полдюжины разных слоев. В данном случае слои имеет глазурь для пирога, но не сам пирог. Как в помещении для больших телефонных станций, провода повсюду поднимаются из пола и проходят вверх через все эти слои коры мозга. Другие провода уходят вниз , в глубину белого вещества. Или просто в сторону. Дендриты каждого нейрона также поднимаются вверх, через несколько слоев, соединяясь с различными источниками сигналов от других нейронов. Каждый нейрон является узлом, где логически сходятся тысячи таких проводов, где входящие сообщения обрабатываются и, иногда, превращаются в исходящие сообщения. Каждый нейрон работает подобно маленькому компьютеру, учитывая влияние тысяч сигналов на своих дендритах и теле клетки, и иногда посылая общее "письмо" для многих своих корреспондентов, через свой длинный и тонкий отросток -- аксон. Иногда, логика соединений нейронов выглядит как "хвост прикрепляется сбоку следующего". Четвертый уровень коры получает большинство входных сигналов от таламуса, расположенного в центре мозга, который, в основном, передаёт сообщения от органов чувств: глаз, ушей, кожи, мускулов,... Нейроны четвертого уровня принимают эти таламусные сообщения и посылают большинство своих импульсов на второй и на третий уровень коры мозга. Некоторые нейроны второго и третьего уровня посылают сообщения вниз, к пятому и шестому уровню. Шестой уровень шлет сообщения назад, в таламус, через белое вещество. И пятый уровень шлет свои сигналы в другие удаленные и/или глубинные нейронные структуры, иногда даже в спинной мозг. Простейший путь сигнала "из глубины" приходит на уровень четыре, затем выше на третий, вниз на пятый или шестой и далее он выходит из коры к какой-нибудь "подкорковой" структуре.
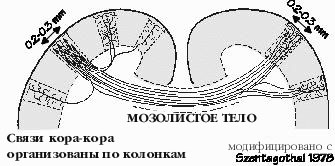
"Это простая диаграмма.", сказал Нейл, посмотрев на свой набросок на салфетке. Он только что вернулся с одного из последних приемов в клинике, после сдачи , как он считает, последнего нейрофизиоло-гического теста. "Так вы уже получили полную схему всех связей в мозге?" У нас нет даже большей ее части. "Похоже, как будто четвертый уровень занимается сортировкой входящей почты." Верно. А глубинные слои специализируются на исходящей почте, однако, без заклейки конвертов и налепливании марок. Верхнии слои производят множество внутриофисных сообщений, часто посылая сообщения в сторону, горизонтально, а не вертикально, по нашей диаграмме. Мы называем их связями типа кора-кора. "Иногда сообщение уходит всего на миллиметр в сторону, а иногда от фронтальной до задней части мозга. Или из левого полушария в правое полушарие, минуя мозолистое тело, самый большой "кабельный канал" мозга. Часто эти внутри-корковые сообщения рассылаются по определенному алгоритму, как будто почта рассылает их в каждый первый дом квартала, но не рассылает их остальным. "Это то что я называю селективной массовой рассылкой." И это происходит если почта исходит из первого дома в квартале. Третий дом в квартале шлет почту третьему дому в другом квартале и, возможно, некоторым другим таким домам, в других кварталах. "Каких размеров этот "квартал" ?" Около половины миллиметра, но он не отделяется от других "перекрестками улиц", только промежутком между адресатами. Сообщение может также следовать через весь город, по "проводам", которые погружаются в белое вещество и потом поднимаются на некотором расстоянии обратно в кору. Рассылка от одного усредненного нейрона затрагивает менее одного процента нейронов, отстоящих от него менее чем на миллиметр.
Вместе с тем, под каждым квадратным миллиметром
коры располагаются 148 000 нейронов, около ста миллионов на квадратном
дюйме. Они часто организованы в миниколонки, около ста нейронов в каждой.
Миниколонки, в свою очередь, иногда организуются в макроколонки из,
приблизительно, 300 миниколонок с 30,000-ми нейронов.
Шесть слоев коры в разных зонах различаются
по толщине. Но, при этом, общая толщина коры остается практически неизменной
для всего мозга. Представь себе город, задания которого ограничины в
высоту шестью этажами. Но высота потолка на этажах не одинакова. Ты
можешь найти здания у которых первый этаж гораздо выше остальных. В
одной части города здания были зачем-то построены с очень высоким третьим
этажом, но потолок на четвертом настолько низок, что стукаешься головой.
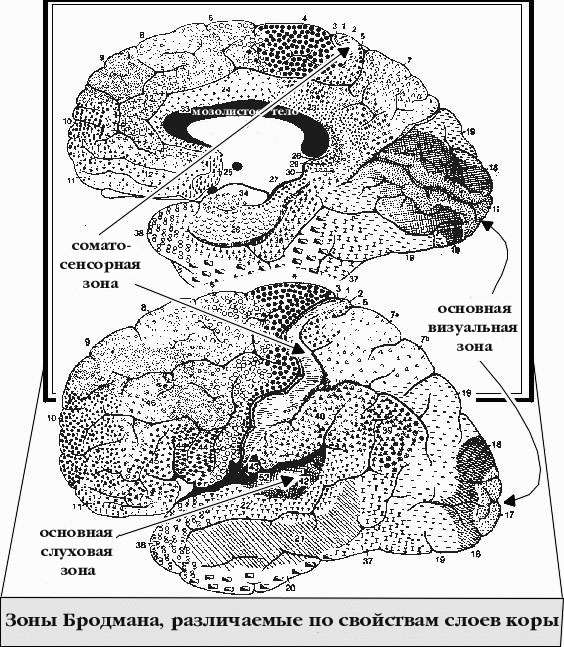
В начале двадцатого века нейроанатомисты определили 52 зоны коры головного
мозга человека, расклассифицировав их по размеру нейронов и относительную
толщину нейронных слоев. Всем зонам присвоены номера. Зона номер 17,
например, известна еще как основная визуальная зона.
|

|
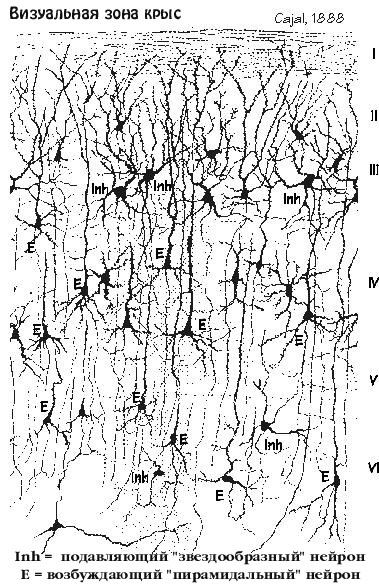
КОГДА ЗИГМУНД ФРЕЙД много лет назад смотрел на мозг
в микроскоп, серое вещество вызлядело как плотная, запутанная паутина,
пересекающихся аксонов, Казалось что все соединяется со всем. В некоторых
пересечениях, вместо пойманных мух, находились темные пузырьки, -- клетки.
Нигде никаких стрелок, ни надписей, ни схем, как здесь перемещается
информация. Возможно Фрейд был скорее разачарован такой картиной, и,
пока техника не улучшила разрешающей способности микрскопов, он начал
изучать мозг с совсем другой точки зрения - психоанализом.
|
|
|
|
ПРЕСЕЧЕНИЕ НЕЙРОННОЙ СВЯЗИ является более классическим
способом влияния на поток информации в мозге. Локальные анестетики могут
блокировать воспроизведение импульса посреди аксона. Парализующие вещества
обычно прерывают работу синапса между нервом и мускульными волокнами.
Некоторые вещества, такие как кураре, затыкают рецепторы, контролирующие
постсинаптические поры так, что ацетохолин не может с ними связаться.
Другие вещества останавливают переработку нейромедиатора после импульса
и, значит, недопускают следующие импульсы. |
|
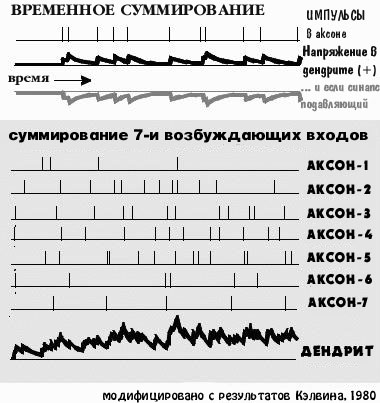
СЛОВА ВОЗБУЖДЕНИЕ И ПОДАВЛЕНИЕ имеют множество значенй,
даже Фрейдистские, но для нейрофизиологов они означают что-то близкое
к сложению и вычитанию, или к вложению денег и их изьятию. Нейрон не
вычитает одну молекулу подавляющего неромедиатора из двух молекул
возбуждающего нейромедиатора. Он избегает проблем с поштучным сложением
чего-либо, автоматически складывая отрицательный или положительный ток,
производимые нейромедиаторами.
[звуковой файл RealAudio
(около 50k)]
|
Отредактировано в ноябре 1996-го. Перевод - 1999 г.